Speciazione
La speciazione riguarda la formazione delle specie. È una parte importante della biologia evolutiva. Darwin pensava che la maggior parte delle specie sorgesse direttamente da specie preesistenti. Questo si chiama anagenesi: specie per cambiament…
La speciazione riguarda la formazione delle specie. È una parte importante della biologia evolutiva.
Darwin pensava che la maggior parte delle specie sorgesse direttamente da specie preesistenti. Questo si chiama anagenesi: specie per cambiamento, o 'evoluzione fletica'. Per gran parte del 20° secolo si è pensato che la maggior parte delle specie sorgesse dalla scissione di specie precedenti: cladogenesi. L'opinione generale era che la maggior parte delle scissioni di specie fosse causata o aiutata nel suo percorso da meccanismi di isolamento.
Senza dubbio la separazione fisica di specie che una volta vivevano insieme è un fattore principale. È illustrato da tanti esempi, alcuni dei quali sono discussi di seguito.
Tuttavia, il lavoro degli ultimi 20 anni ha mostrato altre cause. L'analisi della sequenza del DNA degli esseri viventi ha dimostrato che spesso c'è una certa ibridazione tra specie affini. Ciò significa che i geni sono stati trasferiti da questi incroci. A sua volta, questo significa che l'isolamento riproduttivo non è l'unica definizione di una specie, e la speciazione non ha sempre bisogno di allopatria (le specie devono essere separate riproduttivamente). Le sezioni seguenti illustrano l'idea che la separazione fisica sia stata di primaria importanza nella formazione di nuove specie.
Galleria di immagini
10 Immagini









Meccanismi di isolamento
I meccanismi di isolamento sono cose che impediscono la riproduzione di successo tra gruppi in una specie. L'isolamento riproduttivo delle popolazioni è stabilito. Questo è particolarmente importante per il concetto di specie biologica, poiché le specie sono definite dall'isolamento riproduttivo.
I meccanismi di isolamento possono essere divisi in due gruppi, prima e dopo la fecondazione.
Prima della fecondazione
Fattori che impediscono agli individui di accoppiarsi.
- Isolamento geografico: Le specie si trovano in aree diverse e sono spesso separate da barriere.
- Isolamento temporale: Gli individui non si accoppiano perché sono attivi in momenti diversi. Può trattarsi di diverse ore del giorno o di diverse stagioni. I periodi di accoppiamento delle specie possono non coincidere. Gli individui non si incontrano l'un l'altro durante i loro periodi di accoppiamento, o per niente.
- Isolamento ecologico: Gli individui si accoppiano solo nel loro habitat preferito. Non incontrano individui di altre specie con preferenze ecologiche diverse.
- Isolamento comportamentale: Individui di specie diverse possono incontrarsi, ma non si riconoscono i segnali sessuali che possono essere dati. Un individuo sceglie un membro della propria specie nella maggior parte dei casi.
- Isolamento meccanico: La copulazione può essere tentata ma il trasferimento dello sperma non avviene. Gli individui possono essere incompatibili a causa delle dimensioni o della morfologia.
- Incompatibilità gametica: Il trasferimento dello sperma ha luogo, ma l'uovo non viene fecondato.
Dopo la fecondazione
Fattori che impediscono il successo dell'accoppiamento, come l'incompatibilità genetica, l'invivibilità dell'ibrido o la sterilità.
- Mortalità zigotica: L'uovo viene fecondato, ma lo zigote non si sviluppa.
- Invalidità dell'ibrido: l'embrione ibrido si forma, ma non è vitale.
- Sterilità ibrida: l'ibrido è vitale, ma l'adulto risultante è sterile.
- Ripartizione degli ibridi: Gli ibridi di prima generazione (F1) sono vitali e fertili, ma le generazioni successive di ibridi (F2 e backcross) sono invincibili o sterili.

Isolamento geografico
Si pensa che questa sia la causa più comune della speciazione. La prima persona a pensarci fu Moritz Wagner, un esploratore e storico naturale tedesco.
La prima carriera di Wagner fu quella di geografo, e pubblicò una serie di libri geografici sul Nord Africa, il Medio Oriente e l'America Tropicale. Era anche un appassionato naturalista e collezionista, ed è per questo lavoro che è meglio conosciuto tra i biologi. Ernst Mayr, l'evoluzionista e storico della biologia, ha dato un resoconto dell'importanza di Wagner. p562-565
Durante i suoi tre anni in Algeria, Wagner (tra le altre attività) ha studiato i coleotteri senza volo Pimelia e Melasoma. Ogni genere è diviso in un certo numero di specie, ognuna delle quali è confinata in un tratto della costa nord tra i fiumi che scendono dalle montagne dell'Atlante al Mediterraneo. Appena si attraversa un fiume, appare una specie diversa ma strettamente legata.
"... una specie incipiente [sorgerà] solo quando alcuni individui trasgrediranno i confini limitanti della loro gamma... la formazione di una nuova razza non riuscirà mai... senza una lunga e continua separazione dei colonizzatori dagli altri membri della loro specie".
Questa era una prima descrizione di un processo di un tipo di speciazione geografica. Nel 1942 fu reintrodotta da Mayr, e l'importanza della speciazione geografica divenne una delle idee centrali della sintesi evolutiva.
Un altro termine per la speciazione geografica è la speciazione allopatrica. Allopatria significa "terra diversa".
Isole vacanti
Le isole vulcaniche si formano senza vita, e tutta la vita deve arrivare trasportata dal vento o dall'acqua. Sappiamo dalle isole Hawaii e dalle Galapagos che tutte le forme di vita cambiano quando arrivano sulle isole dalla terraferma.
In circa 6.500 miglia quadrate (17.000 km2), le isole Hawaii hanno la più varia collezione di mosche drosofile del mondo, che vivono dalle foreste pluviali ai prati di montagna. Si conoscono circa 800 specie di drosofilidi hawaiani.
Gli studi mostrano un chiaro "flusso" di specie dalle vecchie alle nuove isole. Ci sono anche casi di colonizzazione verso le isole più vecchie, e di salto di isole, ma questi sono molto meno frequenti.
Con la datazione radioattiva al potassio/argon, le isole attuali sono datate da 0,4 milioni di anni fa (mya) (Mauna Kea) a 10mya (Necker). Il membro più antico dell'arcipelago hawaiano ancora sopra il mare è l'atollo di Kure, che può essere datato a 30 mya.
L'arcipelago stesso, prodotto dal movimento della placca del Pacifico su un punto caldo, esiste da molto più tempo, almeno fino al Cretaceo. Le isole Hawaiane più le isole precedenti che sono ora sotto il mare formano la catena Hawaiian-Emperor seamount; e molte delle montagne sottomarine sono guyot.
Tutte le specie native di drosofilidi nelle Hawaii sono apparentemente discendenti da una singola specie ancestrale che ha colonizzato le isole, circa 20 milioni di anni fa. La successiva radiazione adattativa è stata stimolata dalla mancanza di concorrenza e da un'ampia varietà di nicchie libere. Anche se sarebbe possibile che una singola femmina incinta colonizzasse un'isola, è più probabile che sia stato un gruppo della stessa specie.
Ci sono altri animali e piante nell'arcipelago hawaiano che hanno subito simili, anche se meno spettacolari, radiazioni adattative.
Specie di anello
In biologia, una specie ad anello è una serie collegata di popolazioni vicine, ognuna delle quali può incrociarsi con le popolazioni vicine. Le due estremità della catena si sovrappongono.
Le due popolazioni finali della serie sono troppo lontane per incrociarsi. Tali popolazioni "finali" non consanguinee, anche se geneticamente connesse, possono coesistere nella stessa regione, chiudendo così un "anello".
Le specie ad anello forniscono un'importante prova dell'evoluzione: illustrano ciò che accade nel tempo quando le popolazioni divergono geneticamente. Richard Dawkins ha osservato che le specie ad anello "ci mostrano solo nella dimensione spaziale qualcosa che deve sempre accadere nella dimensione temporale".
Tuttavia, è difficile trovare un esempio semplice e diretto.
Larus gabbiani
Un esempio classico di specie ad anello è l'anello circumpolare dei gabbiani Larus. La gamma di questi gabbiani forma un anello intorno al Polo Nord, che normalmente non viene sorvolato dai singoli gabbiani.
In generale, i gabbiani possono incrociarsi in una certa misura con i loro vicini, tranne alle due estremità dell'anello.
I gabbiani neri minori e i gabbiani reali sono sufficientemente diversi da non ibridarsi normalmente; così il gruppo di gabbiani forma un continuum tranne dove i due lignaggi si incontrano in Europa.
Anche se la realtà è molto più complicata, mostra bene l'idea generale.
Salamandre Ensatina
La salamandra Ensatina è una specie ad anello nelle montagne intorno alla Central Valley californiana. Il complesso forma un ferro di cavallo intorno alle montagne. Anche se l'incrocio può avvenire tra ciascuna delle 19 popolazioni intorno al ferro di cavallo, la sottospecie Ensatina eschscholtzii all'estremità occidentale del ferro di cavallo non può incrociarsi con l'Ensatina klauberi all'estremità orientale. È un'illustrazione di "quasi tutte le fasi di un processo di speciazione" (Dobzhansky). Richard Highton ha sostenuto che l'Ensatina è un caso di specie multiple e non un continuum di una specie.
Il Fringuello verdastro
Il Fringuello verdastro (Phylloscopus trochiloides) ha un certo numero di sottospecie, di cui P. t . viridianus è il più familiare in Europa. Si tratta di una specie ad anello con popolazioni che divergono ad est e ad ovest dell'altopiano tibetano, incontrandosi poi sul lato nord. Le loro relazioni sono abbastanza confuse.




Speciazione simpatrica
La speciazione simpatrica si riferisce alla formazione di due o più specie discendenti da una singola specie ancestrale che occupa la stessa posizione geografica. Oggi si pensa che questo sia molto comune.
Nella speciazione simpatrica, le specie divergono mentre abitano lo stesso luogo. Esempi spesso citati di speciazione simpatrica si trovano negli insetti che diventano dipendenti da diverse piante ospiti nella stessa area.
L'esistenza della speciazione simpatrica come meccanismo di speciazione è stata fortemente contestata. Le persone hanno sostenuto che le prove di speciazione simpatrica sono in realtà esempi di speciazione microgeografica. In generale, questa non è ora la spiegazione preferita. Piuttosto, è il risultato dell'ibridazione tra specie strettamente correlate, seguita dalla selezione naturale che lavora sulla prole di tali incroci. Un esempio ampiamente accettato di speciazione simpatrica è quello dei ciclidi del lago Nabugabo in Africa orientale, che si pensa sia dovuto alla selezione sessuale.
Speciazione tramite poliploidizzazione
La poliploidia ha causato molti eventi di speciazione rapida perché la prole, per esempio, di accoppiamenti tetraploidi x diploidi spesso risulta in progenie sterile triploide.
Tuttavia, non tutti i poliploidi sono riproduttivamente isolati dalle loro piante parentali, e il flusso genico può ancora verificarsi, ad esempio attraverso accoppiamenti ibrido triploide x diploide che producono tetraploidi.
Molte delle piante esistenti e la maggior parte delle specie animali hanno apparentemente subito una poliploidizzazione nella loro storia evolutiva. La riproduzione delle specie poliploidi di successo è talvolta asessuata, per partenogenesi. Per ragioni sconosciute molti organismi asessuati sono poliploidi.
Mosca di biancospino
Un esempio di evoluzione al lavoro è il caso della mosca del biancospino, Rhagoletis pomonella, che sembra subire una speciazione simpatrica.
Diverse popolazioni di mosca del biancospino si nutrono di frutti diversi. Una popolazione distinta è emersa in Nord America nel 19° secolo qualche tempo dopo l'introduzione delle mele, una specie non nativa. Questa popolazione che si nutre di mele si nutre normalmente solo di mele e non del frutto storicamente preferito del biancospino. L'attuale popolazione che si nutre di biancospino non si nutre normalmente di mele.
Alcune prove suggeriscono che la speciazione simpatrica si sta verificando. Le mosche del biancospino maturano più tardi nella stagione e impiegano più tempo a maturare rispetto alle mosche del melo; e ci sono poche prove di ibridazione (i ricercatori hanno documentato un tasso di ibridazione del 4-6%).
L'emergere della nuova mosca del biancospino è un esempio di evoluzione in corso.
Ibridazione
Raramente, una nuova specie si forma quando membri individuali di specie diverse si accoppiano. Di solito, i prodotti di tali accoppiamenti ibridi sono sterili (non fertili), o relativamente tali, e quindi vengono eliminati dalla selezione naturale.
È stato trovato un esempio di una nuova specie ibrida di successo. La nuova specie è un ibrido dei passeri italiani e spagnoli, e i fatti di base sono stati controllati dall'analisi della sequenza del DNA nel loro sangue. L'uccello vive in Italia in una regione dove vivono entrambe le specie madri. Non si riproduce con il passero spagnolo anche se vive con loro fianco a fianco.
Speciazione artificiale
Nuove specie sono state create dall'allevamento di animali domestici, ma le date e i metodi iniziali non sono chiari. Per esempio, le pecore domestiche sono state create per ibridazione, e non producono più prole vitale con Ovis orientalis, una specie da cui discendono.
Il bestiame domestico, d'altra parte, può essere considerato la stessa specie di diverse varietà di bue selvatico, gaur, yak, ecc.
Specie di laboratorio
Le creazioni meglio documentate di nuove specie in laboratorio sono state eseguite alla fine degli anni '80. William Rice e G.W. Salt hanno allevato moscerini della frutta, Drosophila melanogaster, usando un labirinto con tre diverse scelte di habitat come luce/buio e umido/asciutto.
Ogni generazione veniva collocata nel labirinto, e i gruppi di mosche che uscivano da due delle otto uscite venivano separati per riprodursi tra loro nei rispettivi gruppi. Dopo trentacinque generazioni, i due gruppi e la loro prole furono isolati riproduttivamente a causa delle loro forti preferenze di habitat: si accoppiavano solo all'interno delle aree che preferivano, e quindi non si accoppiavano con le mosche che preferivano le altre aree. La storia di questi tentativi è descritta in Rice e Hostert (1993).
Diane Dodd è stata anche in grado di mostrare come l'isolamento riproduttivo può svilupparsi dalle preferenze di accoppiamento in Drosophila pseudoobscura dopo solo otto generazioni utilizzando diversi tipi di cibo, amido e maltosio.
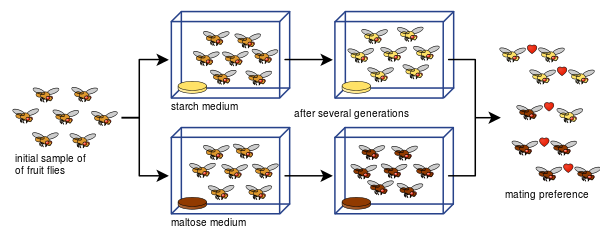
L'esperimento di Dodd è stato facile da replicare per molti altri, anche con altri tipi di moscerini della frutta e alimenti.
Rinforzo (effetto Wallace)
Il rinforzo è il processo con cui la selezione naturale aumenta l'isolamento riproduttivo.
Può verificarsi dopo che due popolazioni della stessa specie sono separate e poi tornano in contatto. Se il loro isolamento riproduttivo era completo, allora si saranno già sviluppate in due specie separate e incompatibili.
Se il loro isolamento riproduttivo è incompleto, allora ulteriori accoppiamenti tra le popolazioni produrranno ibridi, che possono essere fertili o meno. Se gli ibridi sono sterili, o fertili ma meno adatti dei loro antenati, allora non ci sarà un ulteriore isolamento riproduttivo e la speciazione è essenzialmente avvenuta (ad esempio, come nei cavalli e negli asini).
Il ragionamento dietro a questo è che se i genitori della prole ibrida hanno ciascuno tratti selezionati naturalmente per i loro diversi ambienti, la prole ibrida porterà tratti da entrambi, e non si adatterebbe a nessuna delle due nicchie così bene come i due genitori. La bassa fitness degli ibridi causerebbe la selezione per favorire l'accoppiamento assortito, che ridurrebbe l'ibridazione.
Questo è talvolta chiamato l'effetto Wallace dal nome del biologo evoluzionista Alfred Russel Wallace che ha suggerito alla fine del XIX secolo che potrebbe essere un fattore importante nella speciazione.
Se la prole ibrida è più in forma dei loro antenati, allora le popolazioni si fonderanno di nuovo nella stessa specie nell'area in cui sono in contatto.
Il rinforzo è necessario per la separazione quando c'è una "zona ibrida" tra due forme di una specie. Le zone ibride sono regioni in cui popolazioni divergenti si incontrano e si incrociano. La prole ibrida è molto comune in queste regioni, che di solito sono create da specie divergenti che entrano in contatto secondario.
Senza rinforzo le due popolazioni o specie avrebbero un incrocio incontrollabile. Il rinforzo può essere indotto in esperimenti di selezione artificiale come descritto sopra.
Domande e risposte
D: Che cos'è la speciazione?
R: La speciazione è il processo di formazione delle specie. È una parte importante della biologia evolutiva.
D: Cosa pensava Darwin della formazione delle specie?
R: Darwin pensava che la maggior parte delle specie provenisse direttamente da specie preesistenti, cosa che lui chiamava anagenesi o 'evoluzione fitica'.
D: Come veniva vista la speciazione in generale nel 20° secolo?
R: Nel XX secolo, gli scienziati ritenevano che la maggior parte delle specie si formasse quando le specie precedenti si dividevano, il che era noto come cladogenesi. Si credeva anche che questa scissione fosse causata o aiutata da meccanismi di isolamento, come la separazione fisica.
D: In che modo la ricerca recente ha cambiato la nostra comprensione della speciazione?
R: La ricerca recente ha dimostrato che può verificarsi l'ibridazione tra specie affini e che i geni possono essere trasferiti attraverso questi incroci. Ciò significa che l'isolamento riproduttivo non è sempre necessario per definire una specie e l'allopatria (separazione fisica) potrebbe non essere richiesta per la speciazione.
D: Cosa c'entra la separazione fisica con la formazione di nuove specie?
R: Si ritiene che la separazione fisica tra specie un tempo coesistenti sia un fattore principale nella formazione di nuove specie, illustrato da molti esempi discussi nella letteratura scientifica.
Articoli correlati
Autore
AlegsaOnline.com Speciazione Leandro Alegsa
URL: https://it.alegsaonline.com/art/92570
Fonti
- doi.org : 10.1126/science.23.587.506
- pubmed.ncbi.nlm.nih.gov : 17789700
- doi.org : 10.1086/279001
- science.siu.edu : "The concept of species"
- abacus.gene.ucl.ac.uk : "Isolating mechanisms"
- mallet.oeb.harvard.edu : mallet.oeb.harvard.edu/publications/what-speciation
- books.google.com : "The Continental Drift Controversy: Introduction of Seafloor Spreading," p. 292
- pubs.usgs.gov : Volcanism in Hawaii, U.S. Geological Survey Professional Paper 1350, pp. 5-54
- rspb.royalsocietypublishing.org : "The herring gull complex is not a ring species"
- doi.org : 10.1098/rspb.2004.2679
- journals.royalsoc.ac.uk : Electronic Appendix
- pnas.org : Incipient species formation in salamanders of the Ensatina complex
- jstor.org : "Is Ensatina eschscholtzii a ring-species?"
- doi.org : 10.1146/annurev.genet.34.1.401
- pubmed.ncbi.nlm.nih.gov : 11092833